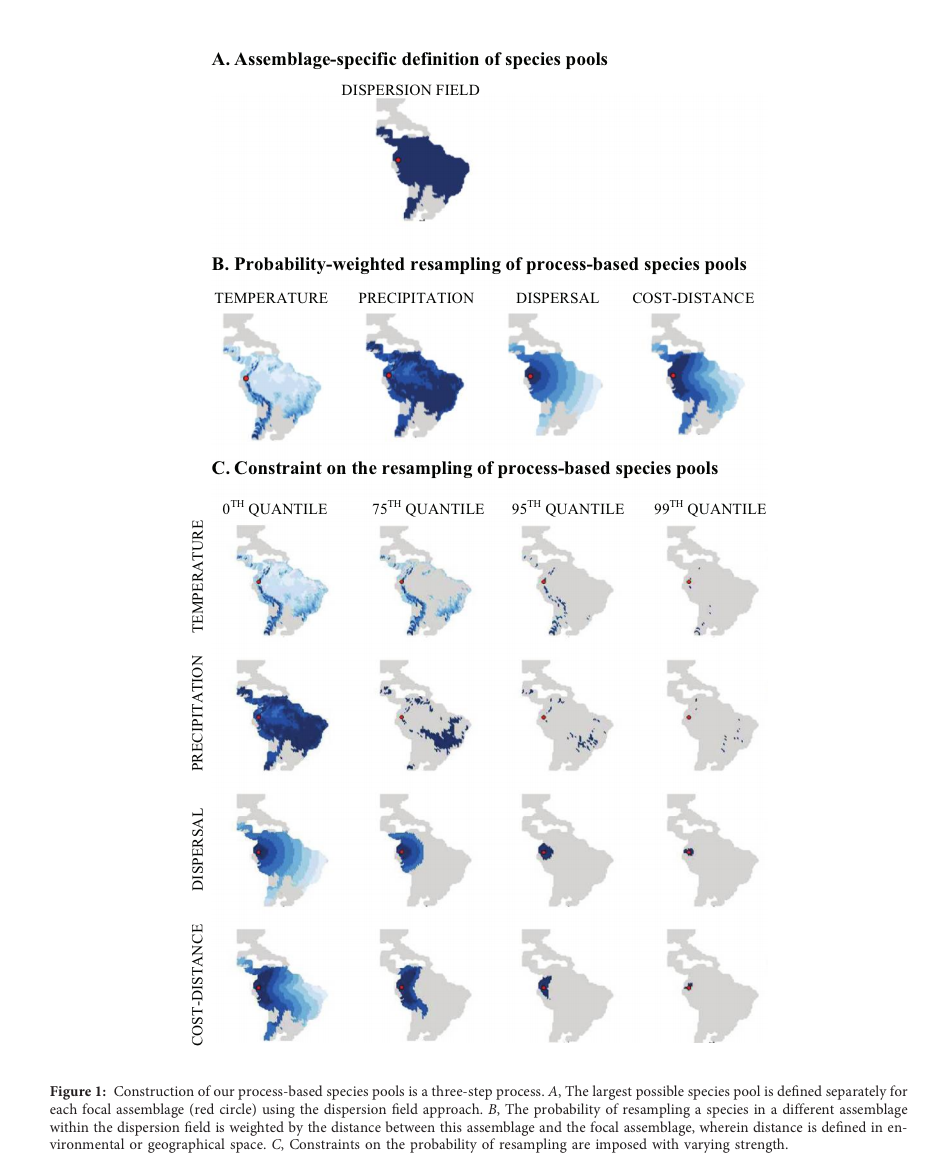
The search for generality is perhaps the greatest driver of modern ecology and probably also the greatest source of ecological angst. Though ecological trends frequently reflect the newest, brightest hope for generality, the search for generality (perhaps by definition) encourages us to ignored details and complexities. Maybe this means that some areas of study won't develop fully until they've fallen out of fashion. And maybe this means that the most interesting science happens when the pressure to 'save community ecology' is gone. A great example of the kind of post-hype, thoughtful approach for trait-based ecology comes from Reynolds et al. (2017) in Tree Physiology. They do a really nice job of highlighting some of the details that must inform trait-based ecology. Here, Reynolds et al. take a broad comparative approach across species, but incorporate important details that have at times been overlooked - especially the role of the environment, recognizing and measuring both constitutive and plastic traits, captures that there are multiple paths (or trait combinations) that can result in similar functioning.
The authors look at four conspecific tree species (Brachychiton spp.) with different average positions along an observed moisture gradient (CMD or climate moisture deficit). Two species occupied drier areas of Australia ('xeric species'), while the other two were found in more moderate areas ('mesic species'). The authors assumed that the different distributions of these species reflect different hydraulic niches. Were species' hydraulic niches associate meaningfully with their traits, specifically those trait associated with drought stress responses. Though these species are closely related--and so huge divergences in form and function might not be expected--the costs and benefits of drought resistance should differ among the species. In dry environments, drought resistance strategies should be more important, and may select for particular traits or sets of traits. Trait states associated with drought conditions include "reduced leaf area, enhanced stomatal control, safer or more efficient xylem, increased tissue water capacitance...and/or deeper root systems " may all be selected for. On the other hand, investment in these traits when water is not limited is often costly, reducing growth and competition. This suggests a meaningful selective regime associated with the CMD gradient and trait values might exist.
One important, but oft-overlooked aspect of trait ecology is that trait values depend on both genes and the environment. Reynold et al. incorporate this fact this by manipulating water availability between drought and control treatments. They measured both constitutive components of trait values – those driven by genetics and expressed regardless of environments – and the plastic or environment-dependent components. For instance, in the presence of prolonged drought, trees might increase root production or change leaf characteristics. In addition to manipulating water availability between treatments, the authors measured nine traits related to morphology and allocation.
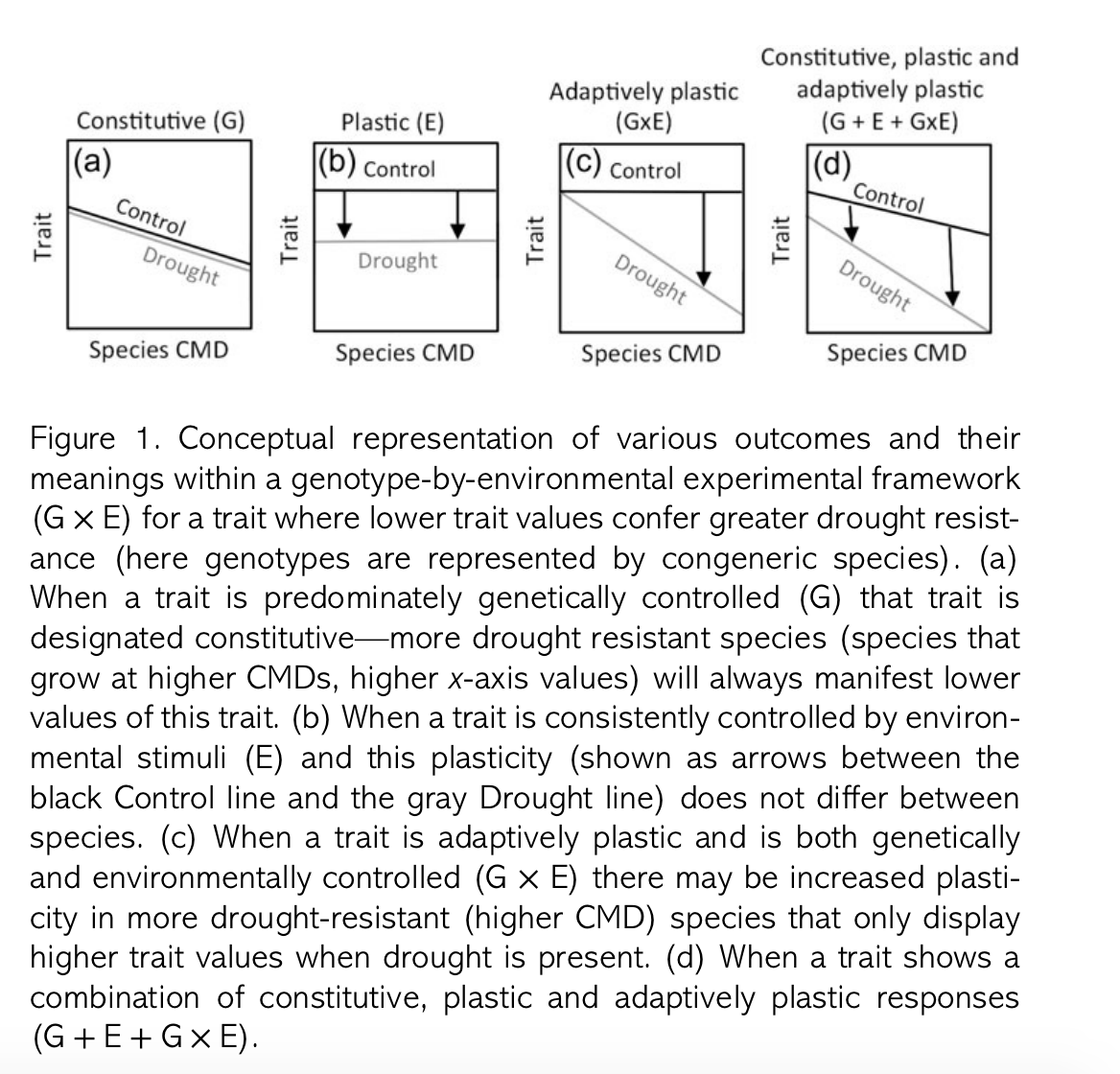 |
| From Reynolds et al. |
Given the expectation that trait values reflect the complex interaction of genetics and the environment in different species, is it possible to even make simple predictions about trait-environment relationships? The authors find that "These complex relationships illustrate that assuming that individual traits (often measured on individuals under a single set of environmental conditions) reflect drought resistance is likely to be overly simplistic and may be erroneous for many species. However, our results do suggest that generalization may be possible, provided multiple traits are measured to explore specific integrated drought strategies."
Indeed, some results are relatively predictable relationships: under well-watered control conditions, the allocation of biomass matched the expectation: xeric species had higher investment in below-ground biomass and in transport tissues than the mesic species (both characteristic of a water-conserving species).
On the other hand, leaf traits such as SLA did not show any trend related to species' assumed drought tolerance, either for constitutive or plastic trait components. Sometimes traits associated with the leaf economic spectrum such as SLA are assumed to indicate stress tolerance, but this was not the case.
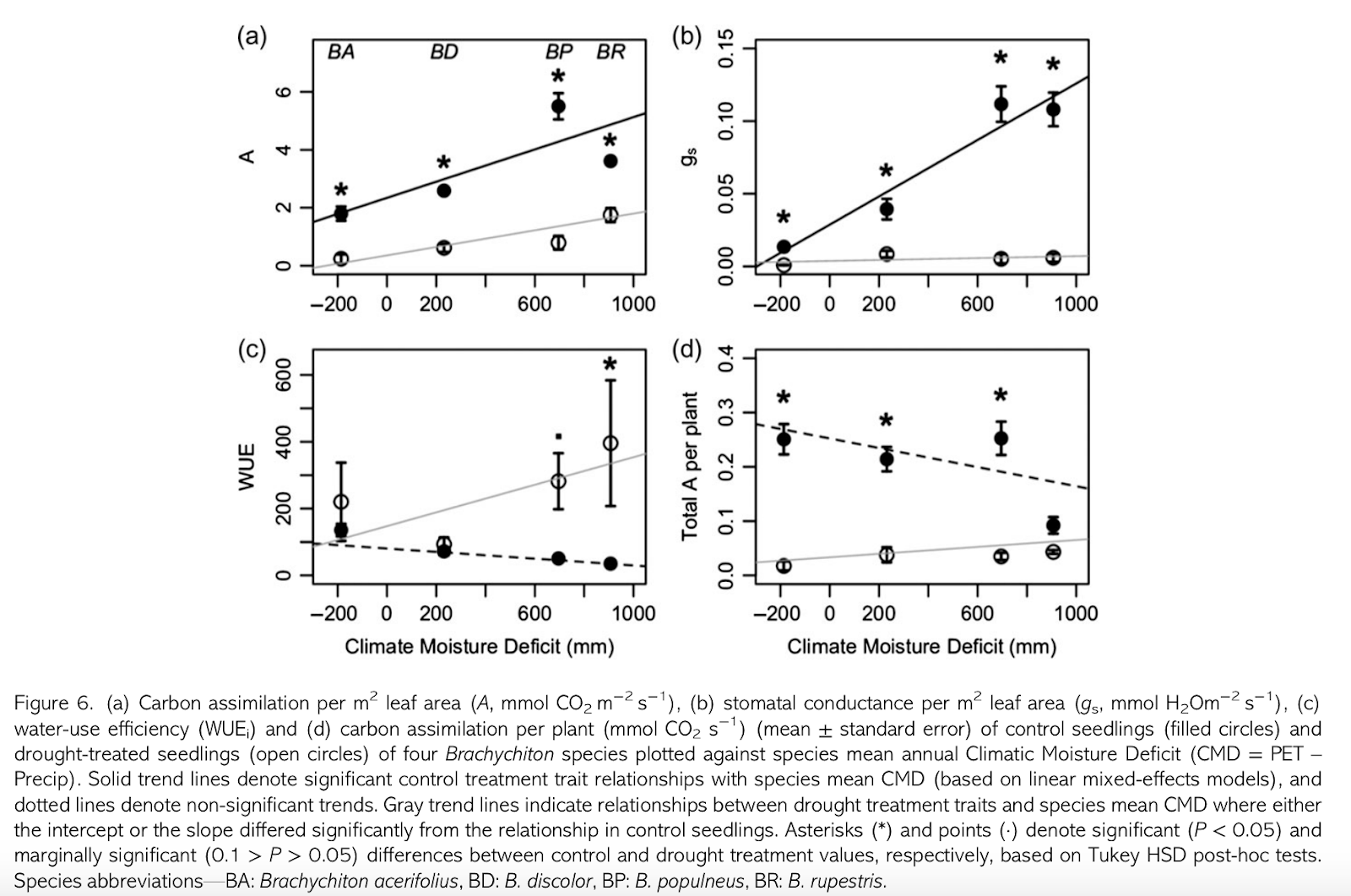
By far the most interesting result was the observation that the xeric species had the highest assimilation and stomatal conductance rate and the lowest water use efficiency under well-watered conditions. Only by also examining these species under drought conditions was it possible to observe that they are highly plastic with regards to water use efficiency. In fact, they show a feast or famine approach to water usage - "where high photosynthetic rates per unit leaf area and high investment in root and stem tissue even in well-watered conditions are achieved through profligate water use during rare periods of water availability, in order to establish a root system and stem storage tissues necessary to survive long periods of water stress." Under drought conditions, these species show reduced root tissue investment; in contrast, mesic species follow expected patterns and plastically increase root tissue investment.
This paper is a reminder that the details are also fascinating and informative. As humans, we may have a simplistic understanding of the realized environment sometimes. To us perhaps all water stress is similar, but for each species in this study, the long term selective pressures may be meaningfully different - in timing, duration, and life stage. This creates the potential for complex differences between species which may best be reflected via life history strategies involving multiple traits. That may still imply some degree of generality is possible, but it is multi-dimensional.
Works cited:
Victoria A Reynolds, Leander D L Anderegg, Xingwen Loy, Janneke HilleRisLambers, Margaret M Mayfield; Unexpected drought resistance strategies in seedlings of four Brachychiton species, Tree Physiology, https://doi.org/10.1093/treephys/tpx143
Victoria A Reynolds, Leander D L Anderegg, Xingwen Loy, Janneke HilleRisLambers, Margaret M Mayfield; Unexpected drought resistance strategies in seedlings of four Brachychiton species, Tree Physiology, https://doi.org/10.1093/treephys/tpx143